

L’étude des espèces actuelles désigne assez clairement la place de l’humanité dans le vivant : juste aux côtés des chimpanzés et des bonobos. Toutefois, cela ne nous dit pas grand-chose sur nos premiers représentants, sur leur biologie et sur leur distribution géographique – bref, sur la manière dont nous sommes devenus humains. Pour cela, il faut essentiellement compter sur la morphologie de trop rares fossiles, les informations paléogénétiques n’étant conservées que pour des périodes récentes – et encore sous des climats plutôt frais !
Ainsi, depuis les années 1960-1970 et la mise en évidence de l’âge très ancien des australopithèques – incluant la fameuse Lucy âgée de 3,18 Ma, découverte en 1974 en Éthiopie – l’acquisition de la bipédie est considérée comme une étape déterminante de l’évolution humaine, une caractéristique essentielle qui marquerait le passage du non-humain à l’humain bien avant le fort accroissement de notre cerveau.
Notre étude, publiée aujourd’hui 24 août dans Nature, des restes du squelette de Sahelanthropus tchadensis, candidat au titre de plus ancien représentant connu de l’humanité, était donc attendue. Alors, bipède ou pas bipède – sous-entendu, humain ou pas humain ? En réalité, poser la question dans ces termes frôle le raisonnement circulaire.
Le dernier ancêtre que nous partageons avec les chimpanzés n’ayant pas été découvert, nous ne connaissons pas l’état initial de la locomotion humaine – bipède ou non.
Les premiers représentants de l’humanité étaient-ils bipèdes ?
Les données les plus anciennes dont nous disposions jusqu’ici étaient les os des membres d’Orrorin (6 Ma, Kenya) et Ardipithecus (5,8 Ma-4,2 Ma, Éthiopie), qui pratiquaient un type de bipédie différent de celui des espèces plus récentes. La bipédie n’est finalement pas un caractère invariant de l’humanité, car elle a une histoire au sein de notre histoire ! La bonne question est donc : les premiers représentants de l’humanité étaient-ils bipèdes, et si oui, dans quelle mesure et comment ? C’est à cette question que notre équipe franco-tchadienne souhaitait apporter des éléments en étudiant les restes nettement plus anciens (environ 7 Ma) de Sahelanthropus.
Sahelanthropus a été initialement décrit en 2002 sur la base d’un crâne (surnommé Toumaï) bien conservé quoique déformé par la fossilisation et de quelques autres spécimens cranio-dentaires découverts par la Mission Paléoanthropologique Franco-Tchadienne (fondée et dirigée par Michel Brunet) à Toros-Ménalla dans le désert du Djourab, au Tchad, représentant au moins trois individus. C’est d’abord sur la morphologie des dents, de la face et de l’arrière-crâne que cette espèce a été rapprochée des fossiles humains plus récents.
Les os des membres décrits dans notre article incluent un fémur partiel gauche (os de la cuisse) et deux ulnae gauche et droit (l’ulna, ou cubitus, est avec le radius l’un des deux os de l’avant-bras ; c’est lui qui forme notre coude). Ces os ont été découverts dans la même localité et la même année que ce crâne mais ont été identifiés ultérieurement, en 2004. Ils appartiennent très vraisemblablement à la même espèce que le crâne, car un seul grand primate a été identifié sur près de 13 800 fossiles représentant une centaine de vertébrés différents collectés dans environ 400 localités à Toros-Ménalla. Par contre, on ne sait pas si ce fémur, ces ulnae et le crâne appartiennent au même individu, car il y a au moins trois individus différents découverts sur ce site.
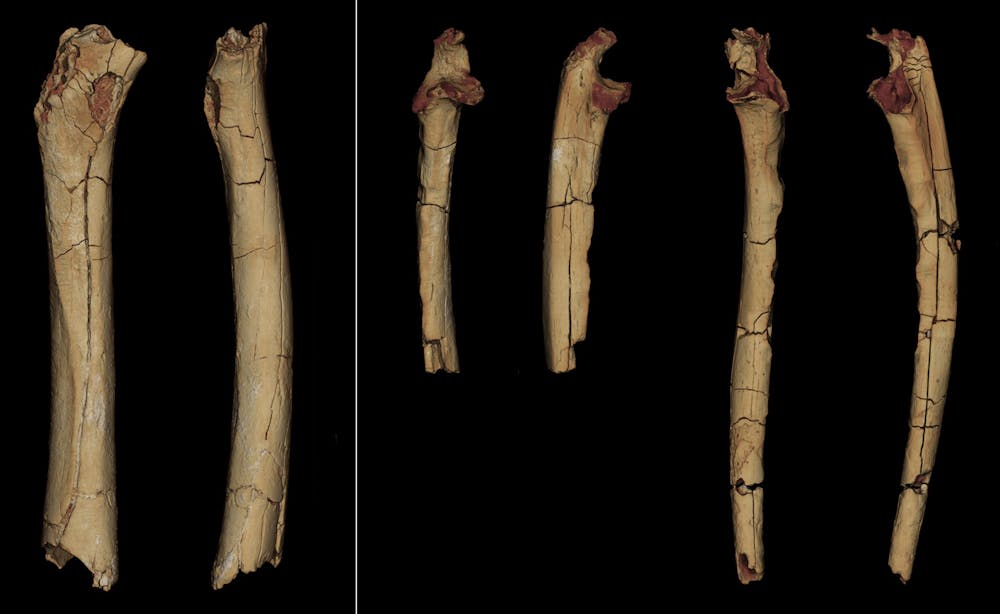
L’analyse de ce matériel, débutée en 2004, a été considérablement ralentie pour diverses raisons, incluant notamment la priorité donnée aux recherches de terrain d’autres restes postcrâniens et à d’autres travaux, ainsi que la difficulté de l’analyse de ce matériel fragmentaire nécessitant des expertises complémentaires. Relancée en 2017, il a fallu cinq années pour la mener à son terme.
Des os étudiés sous toutes les coutures
La préservation de ces os longs n’étant pas très bonne (le fémur, par exemple, a perdu ses deux extrémités), une analyse succincte ne permet pas de fournir des interprétations fiables. Nous les avons donc étudiés sous toutes les coutures, à la fois dans leur morphologie externe et leurs structures internes. Afin de diminuer l’incertitude, nous avons employé diverses approches, incluant des observations directes et des mesures biométriques, ainsi que des analyses d’images 3D des analyses de forme (morphométrie géométrique) et des indicateurs biomécaniques – un travail rarement aussi poussé pour ce type d’étude. Vingt-trois critères ont été comparés à un ensemble de spécimens actuels et fossiles aussi large que possible. Pris séparément, aucun de ces critères ne permet de proposer une interprétation catégorique du matériel – il n’y a pas de caractères « magiques » en paléoanthropologie – et chacun d’entre eux fera certainement l’objet de discussions au sein de la communauté des paléoanthropologues.
Par contre, pris tous ensembles, ces caractères aboutissent à une interprétation de ces fossiles bien plus parcimonieuse que toute autre hypothèse alternative. Cette combinaison indique ainsi que Sahelanthropus pratiquait une bipédie habituelle – c’est-à-dire de manière systématique dans un contexte donné. En l’occurrence, cette bipédie était probablement utilisée pour des déplacements au sol ainsi que dans les arbres. Dans ce dernier cas, elle était très probablement accompagnée d’un autre mode de déplacement : une quadrupédie assurée par des prises fermes de la main qui permettent de saisir les branches, différant clairement de la quadrupédie pratiquée par les gorilles et les chimpanzés qui prennent appui sur le dos de leurs phalanges (« knuckle walking »).
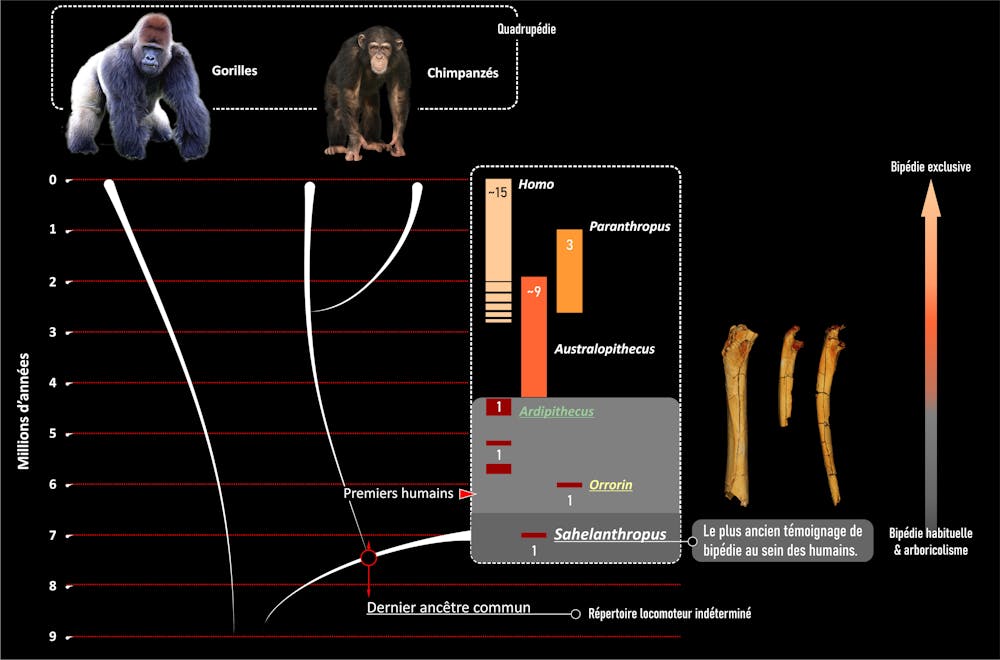
Ces résultats convergents avec les observations menées sur Orrorin et Ardipithecus ont plusieurs implications. Tout d’abord, ils renforcent le concept d’une forme de bipédie très précoce dans l’histoire de l’humanité coexistant avec d’autres modes de locomotion. Il n’y a donc pas eu d’apparition soudaine d’une caractéristique « propre » à l’humanité depuis ses débuts, mais une longue et lente transition sur des millions d’années. Au temps pour le mythe fondateur censé nous démarquer du reste de la biodiversité ! Au contraire, cette phase de l’évolution humaine s’est déroulée selon des modalités tout à fait communes au cours de l’histoire du vivant et du globe, rappelant que l’humanité est un fragment de la biodiversité – un constat qui devrait nous amener à repenser notre attitude envers le monde vivant et les paramètres qui régissent l’hospitalité de notre planète.
Sahelanthropus, Orrorin et Ardipithecus suggèrent également par leurs caractères que l’ancêtre que nous partageons avec les chimpanzés ne ressemblait ni à ces derniers, ni aux bipèdes exclusifs que nous sommes devenus. Contrairement à l’hypothèse d’une conservation de la morphologie ancestrale par les chimpanzés et les bonobos, leur combinaison particulière de grimper vertical et de knuckle walking a plus probablement évolué bien après notre divergence.
Enfin, si Sahelanthropus tchadensis est un témoin de la diversité humaine parmi d’autres, elle est à ce jour la seule espèce bipède habituelle avérée connue à cet âge. En considérant l’ensemble du registre fossile hominoïde faiblement diversifié d’Afrique et d’Eurasie à la fin du Miocène (après 10 Ma), l’acquisition de la bipédie par le rameau humain sur le continent africain reste à ce jour la seule hypothèse bien documentée. Cette bipédie semble à ce stade faire partie d’un répertoire locomoteur opportuniste (flexible, pouvant tirer parti de différents milieux), ce qui correspond bien au paléoenvironnement diversifié de Toros-Ménalla tel que reconstitué par les géologues, paléobotanistes et paléontologues de notre équipe.
[Plus de 80 000 lecteurs font confiance à la newsletter de The Conversation pour mieux comprendre les grands enjeux du monde. Abonnez-vous aujourd’hui]
Ce travail a été développé grâce à une collaboration scientifique nord-sud forte en matière de paléoanthropologie, en l’occurrence entre le laboratoire PALEVOPRIM, le département de paléontologie de l’Université de N’Djamena et le Centre National de Recherche pour le Développement. Alors que ces trois os, appartenant au patrimoine tchadien, vont prochainement retourner dans leur pays, cette collaboration fructueuse va se poursuivre par de nouvelles études de ce matériel mais également de nouvelles recherches sur le terrain, sur les traces du très regretté Yves Coppens, pionnier des recherches paléontologiques au Tchad.
Cet article est republié à partir de The Conversation sous licence Creative Commons. Lire l’article original.